北方和温带地区的多年生树木随着季节变换,表现出显著的生长与休眠周期性转换的生长特性,又叫树木物候,其主要受到光周期和温度的精细调控。在冬季到来之前,树木的顶芽经历连续的适应反应,包括生长停止、驻芽和进入休眠。前人研究发现,短日照介导的杨树顶芽生长停止和建立休眠受红光/远红光受体光敏色素(phyB)及其互作因子PIF8蛋白的调节。然而,位于phyB-PIF8模块下游直接调控杨树在短日照条件下生长停止和进入休眠的分子元件或调控通路仍不清楚。
近日,西南大学生命科学学院罗克明教授团队在国际主流植物学期刊New Phytologist在线发表了题为“SPL16 and SPL23 mediate photoperiodic control of seasonal growth inPopulustrees”的研究论文,报道了短日照条件下PIF8-miR156-SPL16/23-FT2/BRC1级联通路介导调控杨树生长停止和进入休眠过程。该研究建立了光周期感知模块phyB-PIF8与树木物候调控新模块miR156-SPL16/23的直接联系,为树木季节性生长背后的分子机制提供了新的见解。

New Phytologist (2023) doi: 10.1111/nph.19485
本研究对野生型(WT)毛白杨(Populus tomentosa)进行不用光周期处理,发现短日照条件(SD)抑制杨树叶片和顶芽中miR156的表达,而miR156靶向的18个SPL基因中,有11个表达上调,包括SPL16和SPL23(为拟南芥开花促进因子SPL3的同源基因)。相较于WT植株,杨树MIR156a-OE和MIR156c-OE超表达植株在短日照条件下生长停止和驻芽时间均延缓。进一步研究发现,miR156靶标基因SPL16/23在控制杨树生长停止中发挥关键作用。杨树spl16和spl16/23突变体在短日照条件(SD)下呈现生长停止和驻芽延缓的表型;而超表达SPL16或SPL23导致杨树的顶芽生长停止和驻芽提前。因此,SPL16和SPL23是介导短日照诱导杨树生长停止和驻芽的正调控因子(图1)。
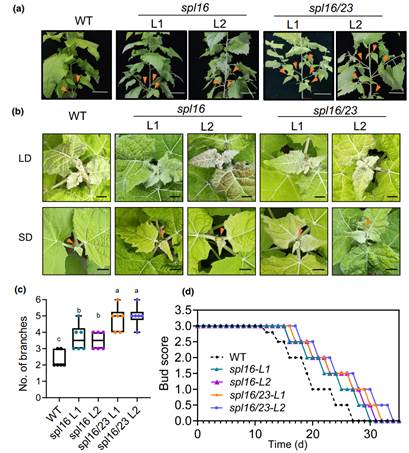
图1.杨树SPL16/23是杨树生长停止和驻芽的正调控因子
为了进一步解析SPL16/23调控杨树季节性生长停止的分子机制,本研究利用Y1H, EMSA, Effector-reporter和DLA等试验证明SPL16/23能直接抑制FT2表达,同时直接促进BRC1.1/1.2表达,进而整合下游通路调控杨树季节性生长停止。此外,本研究揭示miR156-SPL16/23模块作用于phyB-PIF8模块下游发挥作用。PIF8.1和PIF8.2能够直接结合MIR156a/c启动子并抑制其表达,通过上调miR156靶标基因SPL16/23来介导短日照诱导的顶芽生长停止。本研究拓展了miR156-SPL年龄模块在调控多年生树木季节性生长过程中的功能,揭示了短日照条件下PIF8-miR156-SPL16/23-FT2/BRC1级联通路调节杨树生长停止和进入休眠的新机制(图2)。
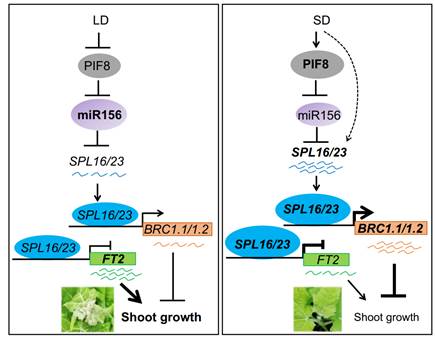
图2. SPL16和SPL23介导光周期调控杨树季节性生长的调控通路
西南大学生命科学学院魏洪彬副教授、硕士研究生罗梦庭、硕士研究生邓娇为论文的共同第一作者。西南大学生命科学学院罗克明教授为论文通讯作者。本项研究得到国家重点研发项目子课题(2022YFD1201600)、国家自然科学基金青年项目(32101483)和重庆市科技兴林重大专项(2D-2022-2)等项目共同资助。

